Комплементарная днк. Днк и гены
Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
ДНК (дезоксирибонуклеиновая кислота) - своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение. ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни как человека, так и любого др. организхма. Искусственное или естественное воздействие внешней среды способны лишь в незначительной степени повлиять на общую выраженность отдельных генетических признаков или сказаться на развитии запрограммированных процессов.
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С ) и фосфатной (Ф ) группы (фосфодиэфирные связи).

Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т ), гуанин — только с цитозином (Г-Ц ). Именно эти пары и составляют «перекладины» винтовой "лестницы" ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
- Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной.
- Двойная спираль «расстегивается» на две ветви, когда ферменты разрушают связь между базовыми парами химических соединений.
- Каждая ветвь является элементом новой ДНК. Новые базовые пары соединяются в той же последовательности, что и в родительской ветви.
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Рис. 4 . Азотистые основания: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты - это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
Нуклеотиды состоят из азотистого основания , соединенного с пятиуглеродным углеводом (пентозой) - дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H 2 PO 3 -).
Азотистые основания бывают двух типов: пиримидиновые основания - урацил (только в РНК), цитозин и тимин, пуриновые основания - аденин и гуанин.

Рис. 5. Типы азотистых оснований: пиримидиновые и пуриновые
Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение выделить 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль . Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей . Аденин всегда соединяется с тимином, а цитозин - с гуанином. Это называется правилом комплементарности .
Правило комплементарности:
| A-T G-C |
Например, если нам дана цепь ДНК, имеющая последовательность
3’- ATGTCCTAGCTGCTCG - 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении - от 5’-конца к 3’-концу:
5’- TACAGGATCGACGAGC- 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ
Репликация ДНК - это процесс удвоения молекулы ДНК путем матричного синтеза. Репликация происходит по полуконсервативному механизму . Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез - это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:
- ДНК-топоизомераза, располагаясь перед вилкой репликации, разрезает ДНК для того, чтобы облегчить ее расплетание и раскручивание.
- ДНК-хеликаза вслед за топоизомеразой влияет на процесс «расплетения» спирали ДНК.
- Связывающие ДНК-белки осуществляют связывание нитей ДНК, а также проводят их стабилизацию, не допуская их прилипания друг к другу.
- ДНК-полимераза синтезирует ведущую цепь дочерней ДНК.

Рис. 9. Схематическое изображение процесса репликации, цифрами отмечены: (1) Запаздывающая нить, (2) Лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) Хеликаза, (10) Белки, связывающие одноцепочечную ДНК, (11) Топоизомераза
Строение РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК, РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Однако в отличие от ДНК, РНК обычно имеет не две цепи, а одну. Пентоза в РНК представлена рибозой, а не дезоксирибозой (у рибозы присутствует дополнительная гидроксильная группа на втором атоме углевода). Наконец, ДНК отличается от РНК по составу азотистых оснований: вместо тимина (Т ) в РНК представлен урацил (U ) , который также комплементарен аденину.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией , то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами.
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК - эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ - 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.
![]()
Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’- ATGTCCTAGCTGCTCG - 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’- TACAGGATCGACGAGC- 3’,
а синтезируемая с нее РНК - последовательность
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка
на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:

Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
ГЕНЕТИЧЕСКИЙ КОД
Генетический код - способ кодирования аминокислотной последовательности белков с помощью последовательности нуклеотидов. Каждая аминокислота кодируется последовательностью из трех нуклеотидов - кодоном или триплетом.
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5" к 3" концу мРНК.
Таблица 1. Стандартный генетический код
|
1-е
ние |
2-е основание |
3-е
ние |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Tyr/Y) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
U C A |
U A A |
Стоп-кодон** |
U G A |
Стоп-кодон** |
A |
||
|
U U G |
U C G |
U A G |
Стоп-кодон** |
U G G |
(Trp/W) |
G |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(His/H) |
C G U |
(Arg/R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
A |
||||
|
A U G |
(Met/M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu/E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
- *Триплет AUG , также кодирующий метионин, называется старт-кодоном . С этого кодона начинается синтез молекулы белка. Таким образом, во время синтеза белка, первой аминокислотой в последовательности всегда будет метионин.
- **Триплеты UAA , UAG и UGA называются стоп-кодонами и не кодируют ни одной аминокислоты. На этих последовательностях синтез белка прекращается.
Свойства генетического кода
1. Триплетность . Каждая аминокислота кодируется последовательностью из трех нуклеотидов - триплетом или кодоном.
2. Непрерывность . Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.
3. Неперекрываемость . Один нуклеотид не может входить одновременно в два триплета.
4. Однозначность . Один кодон может кодировать только одну аминокислоту.

5. Вырожденность . Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность . Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’- CCGATTGCACGTCGATCGTATA - 5’.
Матричная цепь будет иметь последовательность:
5’- GGCTAACGTGCAGCTAGCATAT - 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’- CCGAUUGCACGUCGAUCGUAUA - 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’- AUAUGCUAGCUGCACGUUAGCC - 3’.
Теперь найдем старт-кодон AUG:
5’- AUAUG CUAGCUGCACGUUAGCC - 3’.
Разделим последовательность на триплеты:
звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК - на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
(общие понятия)
Геном - совокупность всех генов организма; его полный хромосомный набор.
Термин "геном" был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими ("избыточными") последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.
В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации ), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент» . Позднее эта концепция была расширена до определения «один ген — один полипептид» , поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
На рис. 14 показана схема того, как триплеты нуклеотидов в ДНК определяют полипептид - аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками(так называемыми интронами ).
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена , кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов ). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
 Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру - нуклеоид. Хромосома прокариота Escherichia coli , чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
См. также: Равин Н.В., Шестаков С.В. Геном прокариот // Вавиловский журнал генетики и селекции, 2013. Т. 17. № 4/2. С. 972-984.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов
|
Общая ДНК, п.н. |
Число хромосом* |
Примерное число генов |
|
|
Escherichia coli (бактерия) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans (нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana (растение) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
20 000 |
|
|
Oryza sativa (рис) |
480 000 000 |
57 000 |
|
|
Mus musculus (мышь) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens (человек) |
3 070 128 600 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам
* Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом. Диплоидный
набор хромосом (от греч. diploos- двойной и eidos- вид) - двойной набор хромосом (2n), каждая из которых имеет себе гомологичную.
**Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
***Для самок с двумя Х хромосомами. У самцов есть Х хромосома, но нет Y, т. е. всего 11 хромосом.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке E. coli (табл. 2). Клетки плодовой мушки Drosophila , классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки E. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2n ) зависит от вида организма (табл. 2).
Например, в соматической клетке человека 46 хромосом (рис. 17 ). Каждая хромосома эукариотической клетки, как показано на рис. 17, а , содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.

Рис. 17. Хромосомы эукариот. а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.
Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или Х и Х), получится последовательность длиной около одного метра. Прим.: У всех млекопитающих и других организмов с гетерогаметным мужским полом, у самок две X-хромосомы (XX), а у самцов — одна X-хромосома и одна Y-хромосома (XY).
Большинство клеток человека , поэтому общая длина ДНК таких клеток около 2м. У взрослого человека примерно 10 14 клеток, таким образом, общая длина всех молекул ДНК составляет 2・10 11 км. Для сравнения, окружность Земли — 4・10 4 км, а расстояние от Земли до Солнца — 1,5・10 8 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК, — это митохондрии и хлоропласты. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл. Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК.
СТРОЕНИЕ ГЕНОВ
Рассмотрим строение гена у прокариот и эукариот, их сходства и различия. Несмотря на то, что ген — это участок ДНК, кодирующий всего один белок или РНК, кроме непосредственно кодирующей части, он также включает в себя регуляторные и иные структурные элементы, имеющие разное строение у прокариот и эукариот.
Кодирующая последовательность - основная структурно-функциональная единица гена, именно в ней находятся триплеты нуклеотидов, кодирующие аминокислотную последовательность. Она начинается со старт-кодона и заканчивается стоп-кодоном.
До и после кодирующей последовательности находятся нетранслируемые 5’- и 3’-последовательности . Они выполняют регуляторные и вспомогательные функции, например, обеспечивают посадку рибосомы на и-РНК.
Нетранслируемые и кодирующая последовательности составлют единицу транскрипции - транскрибируемый участок ДНК, то есть участок ДНК, с которого происходит синтез и-РНК.
Терминатор - нетранскрибируемый участок ДНК в конце гена, на котором останавливается синтез РНК.
В начале гена находится регуляторная область , включающая в себя промотор и оператор .
Промотор - последовательность, с которой связывается полимераза в процессе инициации транскрипции. Оператор - это область, с которой могут связываться специальные белки - репрессоры , которые могут уменьшать активность синтеза РНК с этого гена - иначе говоря, уменьшать его экспрессию .
Строение генов у прокариот
Общий план строения генов у прокариот и эукариот не отличается - и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор. Однако организация генов у прокариот и эукариот отличается.

Рис. 18. Схема строения гена у прокариот (бактерий) - изображение увеличивается
В начале и в конце оперона есть единые регуляторные области для нескольких структурных генов. С транскрибируемого участка оперона считывается одна молекула и-РНК, которая содержит несколько кодирующих последовательностей, в каждой из которых есть свой старт- и стоп-кодон. С каждого из таких участков с интезируется один белок. Таким образом, с одной молекулы и-РНК синтезируется несколько молекул белка.
Для прокариот характерно объединение нескольких генов в единую функциональную единицу - оперон . Работу оперона могут регулировать другие гены, которые могут быть заметно удалены от самого оперона - регуляторы . Белок, транслируемый с этого гена называется репрессор . Он связывается с оператором оперона, регулируя экспрессию сразу всех генов, в нем содержащихся.
Для прокариот также характерно явление сопряжения транскрипции и трансляции .
![]()
Рис. 19 Явление сопряжения транскрипции и трансляции у прокариот - изображение увеличивается
Самовоспроизведение наследственного материала. Репликация ДНК
Репликация – удвоение молекул ДНК, в результате которого образуются две двойные спирали ДНК. Основано на принципах :
1) Комплементарность – каждая из двух цепей – матрица для синтеза комплементарной цепи. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали.
2) Полуконсервативность – каждая из двух двойных спиралей несет одну нить
материнской ДНК.
3) Антипараллельность – каждая цепь ДНК имеет определенную ориентацию: 5"-конец одной цепи соединяется с 3"-концом другой, и наоборот.
4) Прерывистость – репликация осуществляется фрагментами.
Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. origin -начало). Они включают последовательность, состоящую из 300 нуклеотидных пар, узнаваемую специфическими белками. Двойная спираль ДНК в этих локусах разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей - репликационные вилки , которые движутся в противоположных от локуса ori направлениях. Между репликационными вилками образуется структура, называемая репликационным глазком , где на двух цепях материнской ДНК образуются новые полинуклеотидные цепи.
С помощью фермента хеликазы, разрывающего водородные связи, двойная спираль ДНК расплетается в точках начала репликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами, находящимися в нуклеоплазме. На каждой из цепей, образующихся в области репликационной вилки, при участии фермента ДНК-полимеразы осуществляется синтез комплементарных цепей.
В процессе синтеза репликационные вилки движутся вдоль материнской спирали в противоположных направлениях, захватывая все новые зоны.
Разделение спирально закрученных цепей родительской ДНК ферментом хеликазой вызывает появление супервитков перед репликационной вилкой. Это объясняется тем, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, родительская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вся молекула ДНК перед ней должна была бы быстро вращаться, что потребовало бы большой затраты энергии. В действительности это не наблюдается благодаря особому классу белков, называемых ДНК-топоизомеразами . Топоизомераза разрывает одну из цепей ДНК, что дает ей возможность вращаться вокруг второй цепи. Это ослабляет накопившееся напряжение в двойной спирали ДНК.
К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат.
Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3"-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3"-конце.
Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3"-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинуклеотидную цепь называют затравкой или праймером.
Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы . Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3"-ОН-конец.
Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5"- к 3" -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3" → 5") сборка новой цепи происходит непрерывно от 5"- к 3"-концу и она постепенно удлиняется на 3"-конце, то другая цепь, синтезируемая на матрице (5" → 3"), должна была бы расти от 3"- к 5"-концу. Это противоречит направлению действия фермента ДНК-полимеразы.
В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки ) также в направлении от 5"- к 3"-концу (по типу шитья «назад иголкой»). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки.
В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей . Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов,требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5" → 3", в целом эта цепь растет в направлении 3" → 5".
В виду того, что от локуса oriкак правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК.
Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК.
Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: хеликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки.
Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками, что затрудняет ее деспирализацию, необходимую для осуществления репликативного синтеза.
Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации - репликон . Однажды начавшись в точке начала, репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.
В реакции, катализируемой обратной транскриптазой .
кДНК часто используется для клонирования генов эукариот в прокариотах . Комплементарная ДНК также образуется ретровирусами (ВИЧ-1 , ВИЧ-2 , Вирусом иммунодефицита обезьян) и затем интегрируется в ДНК хозяина, образуя провирус .
Часто гены эукариот удается экспрессировать в клетках прокариот. В наиболее простом случае, метод предполагает встраивание эукариотической ДНК в геном прокариот, далее транскрипцию ДНК в мРНК и затем трансляцию мРНК в белки. Клетки прокариот не имеют ферментов для вырезания интронов, и поэтому интроны из ДНК эукариот должны быть вырезаны до момента встраивания в геном прокариот. ДНК, комплементарная зрелой мРНК, таким образом, называется комплементарной ДНК - cDNA (кДНК). Для успешной экспрессии белков, закодированных в эукариотической cDNA в прокариотах, требуются также регуляторные элементы прокариотических генов (например, промоторы).
Одним из методов для получения необходимого гена (молекулы ДНК), которая будет подлежать репликации (клонированию) с выходом значительного количества реплик, является конструирование на мРНК комплементарной относительно неё ДНК (кДНК). Этот метод требует применения обратной транскриптазы - фермента, который присутствует в некоторых РНК-содержимых вирусах и обеспечивает синтез ДНК на РНК матрице.
Метод широко применяется для получения кДНК и включает в себя выделение из тотальной мРНК ткани мРНК, которая кодирует трансляцию определенного белка (например, интерферона, инсулина) с дальнейшим синтезом на этой мРНК как на матрице необходимой кДНК с помощью обратной транскриптазы.
Ген, который был получен с помощью вышеуказанной процедуры (кДНК), необходимо ввести в бактериальную клетку таким образом, чтобы он интегрировался в её геном. Для этого формируют рекомбинантную ДНК, которая состоит из кДНК и особенной молекулы ДНК, которая правит как проводник, или вектор, способный проникать реципиенту в клетку. В роли векторов для кДНК применяют вирусы или плазмиды. Плазмиды - это небольшие кольцевые молекулы ДНК, которые находятся отдельно от нуклеоида бактериальной клетки, содержат в своем составе несколько важных для функции всей клетки генов (например, гены стойкости к антибиотикам и могут реплицироваться независимо от основного генома (ДНК) клетки. Биологически важными и практически полезными для генной инженерии свойствами плазмида являются их способность к переходу из одной клетки в другую по механизму трансформации или конъюгации, а также способность включаться в бактериальную хромосому и реплицироваться вместе с ней.
Принципы решения типовых задач по молекулярной биологии
Задача 1
Одна из цепочек молекулы ДНК имеет следующий порядок нуклеотидов: АЦГ ТАГ ЦТА ГЦГ… Напишите порядок нуклеотидов в комплементарной цепочке ДНК.
Пояснение к решению задачи. Известно что две цепочки в молекуле ДНК соединяются водородными связями между комплементарными нуклеотидами (А-Т, Г-Ц). Порядок нуклеотидов в известной цепочке ДНК:
А Ц Г Т А Г Ц Т А Г Ц Г
Т Г Ц А Т Ц Г А Т Ц Г Ц – порядок нуклеотидов в комплементарной цепочке ДНК.
Ответ: порядок нуклеотидов в комплементарной цепочке ДНК: ТГЦАТЦГАТЦГЦ
Задача 2
Порядок нуклеотидов в одной из цепочек молекулы ДНК следующий: АГЦТАЦГТАЦГА… Определите порядок аминокислот в полипептиде, закодированном этой генетической информацией.
Пояснение к решению задачи . Известно, что матрицей для синтеза полипептида является молекула и-РНК, матрицей для синтеза которой в свою очередь является одна из цепочек молекулы ДНК. Первый этап биосинтеза белка: транскрипция – переписывание порядка нуклеотидов с цепочки ДНК на и-РНК. Синтез и-РНК идет по принципу комплементарности (А-У, Г-Ц). Вместо тимина в и-РНК азатистое основание урацил. Известная цепочка ДНК:
А Г Ц Т А Ц Г Т А Ц Г А…
У Ц Г А У Г Ц А У Г Ц У…-порядок нуклеотидов в и-РНК.
Одну аминокислоту кодируют три рядом расположенные нуклеотида цепочки и-РНК (кодоны) – разбиваем и-РНК на кодоны:
УЦГ, АУГ, ЦАУ, ГЦУ и по тадлице кодонов находим соответствующие им аминокислоты: УЦГ – соответствует серину, АУГ – метионину, ЦАУ – гисцидину, ГЦУ – аланину.
Ответ: порядок аминокислот в закодированном полипептиде: сер – мет – гис – ала…
Задача 3
Одна из цепочек молекулы ДНК имеет следующий порядок нуклеотидов: ГГЦАТГГАТЦАТ…
а) Определите последовательность аминокислот в соответствуюшем полипептиде, если известно, что и РНК синтезируется на комплементарной цепи ДНК.
б) Как изменится первичная структура полипептида, если выпадет третий нуклеотид?
Пояснение к решению задачи .
а) Известно, что молекула и- РНК синтезируется по принципу комплементарности на одной из цепей молекулы ДНК. Нам известен порядок нуклеотидов в одной цепи ДНК и сказано, что и-РНК синтезируется на комплементарной цепи. Следовательно, надо построить комплементарную цепь ДНК, помня при этом, что аденин соответствует тимину, а гуанин цитозину. Двойная цепочка ДНК будет выглядеть следующим образом:
Г Г Ц А Т Г Г А Т Ц А Т…
Ц Ц Г Т А Ц Ц Т А Г Т А …
Теперь можно построить молекулу и – РНК. Следует помнить, что вместо тимина в молекулу РНК входит урацил. Следовательно:
ДНК: Ц Ц Г Т А Ц Ц Т А Г А Т…
И-РНК: Г Г Ц А У Г Г А У Ц А У….
Три рядом расположенных нуклеотида (триплет, кодон) и-РНК определяют присоединение одной аминокислоты. Соответствующие триплетам аминокислоты находим по таблице кодонов. Кодон ГГЦ соответствует гли, А УГ – мет, ГАУ – асп, ЦАУ – гис. Следовательно, последовательность аминокислот участка полипептидной цепи будет: гли – мет – асп – гис…
б) Если в цепочке молекулы ДНК выпадет третий нуклеотид, то она будет выглядеть следующим образом:
ГГАТГГАТЦАТ…
Комплементарная цепочка: ЦЦТАЦЦТАГТА...
Информационная и-РНК: ГГАУГГАУЦАУ...
Произойдут изменения всех кодонов. Первый кодон ГГА соответствует аминокислоте гли, второй УУГ – три, третий АУЦ – иле, четвертый не полный. Таким образом, участок полипептида будет выглядеть:
Гли – три – иле…, т. е. произойдет значительное изменение порядка и количества аминокислот в полипептиде.
Ответ: а) последовательность аминокислот в полипептиде: гли – мет – асп – гис…,
б) после выпадения третьего нуклеотида последовательность аминокислот в полипептиде: гли – три – иле…
Задача 4
Полипептид имеет следующий порядок аминокислот: фен – тре – ала – сер – арг…
а) Определите один из вариантов последовательности нуклеотидов гена, кодирующего данный полипептид.
б) Какие т-РНК (с какими антикодонами) участвуют в синтезе этого белка? Напишите один из возможных вариантов.
Пояснение к решению задачи .
а) Полипептид имеет следующую последовательность аминокислот: фен – тре – ала – сер – арг… По таблице кодонов находим один из триплетов, кодирующий соответствующие аминокислоты. Фен – УУУ, тре – АЦУ, ала – ГЦУ, сер – АГУ, арг – АГА. Следовательно, кодирующая данный полипептид и-РНК будет иметь следующую последовательность нуклеотидов:
УУУАЦУГЦУАГУАГА…
Порядок нуклеотидов в кодирующей цепочке ДНК: АААТГАЦГАТЦАТЦТ…
Комплементарная цепочка ДНК: ТТТАЦТГЦТАГТАГА…
б) По таблице кодонов находим один из вариантов последовательности нуклеотидов и-РНК (как в предыдущем варианте). Антикодоны т-РНК комплементарны кодонам и-РНК:
и-РНК УУУАЦУГЦУАГУАГА…
антикодоны т-РНК ААА, УГА, ЦГА, УЦА, УЦУ
Ответ: а) один из вариантов последовательности нуклеотидов в гене будет:
АААТГАЦГАТЦАТЦТ
ТТТАЦТГЦТАГТАГА,
б) в синтезе этого белка будут участвовать т-РНК с антикодонами (один из вариантов): ААА, УГА, ЦГА, УЦА, УЦУ.
Задачи для самоконтроля
1.Одна из цепочек фрагмента молекулы ДНК имеет такую последовательность нуклеотидов: АГТГАТГТТГГТГТА… Какова будет структура второй цепочки молекулы ДНК?
2.Участок одной цепи ДНК имеет следующую последовательность нуклеотидов: ТГААЦАЦТАГТТАГААТАЦЦА… Какова последовательность аминокислот в полипептиде, соответствующем этой генетической информации?
3.Участок одной нити ДНК имеет такую структуру: ТАТТЦТТТТТГТ… Укажите структуру соответствующей части молекулы белка, синтезированного при участии комплементарной цепи. Как изменится первичная структура фрагмента белка, если выпадет второй от начала нуклеотид?
4.Часть молекулы белка имеет такую последовательность аминокислот: сер – ала – тир – лей – асп… Какие т-РНК (с какими антикодонами) участвуют в синтезе этого белка? Напишите один из возможных вариантов.
5.Напишите один из вариантов последовательности нуклеотидов в гене, если кодируемый белок имеет следующую первичную структуру:
Ала – тре – лиз – асн – сер – глн – глу – асп…
Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом: Соответствие А « Т и Г « Ц называют правилом комплементарности , а сами цепи - комплементарными . Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
ФУНКЦИЯ НУКЛЕИНОВЫХ КИСЛОТ Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация). Репликация и транскрипция . С химической точки зрения синтез нуклеиновой кислоты это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом: Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты ДНК-полимеразы.
Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:
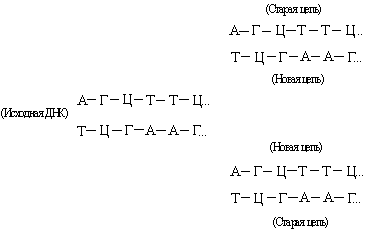 Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации
(см. также
НАСЛЕДСТВЕННОСТЬ)
.
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):
Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации
(см. также
НАСЛЕДСТВЕННОСТЬ)
.
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):
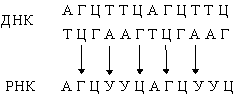 Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК
®
ДНК и ДНК
®
РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК
®
РНК и РНК
®
ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:
Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК
®
ДНК и ДНК
®
РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК
®
РНК и РНК
®
ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:
 Вирусная РНК может транскрибироваться ферментом
-
обратной транскриптазой
-
в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Вирусная РНК может транскрибироваться ферментом
-
обратной транскриптазой
-
в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.